5.2 Sistemas de
reparación.
La reparación del ADN es un conjunto de procesos por los cuales una célula identifica y corrige daños hechos a las moléculas de ADN que codifican el genoma. En las células humanas, tanto las actividades metabólicas como los factores ambientales, como los rayos UV o la radiactividad, pueden causar daños al ADN, provocando hasta un millón de lesiones moleculares por célula por día. Muchas de estas lesiones causan daños estructurales a la molécula de ADN, y pueden alterar o eliminar la capacidad de la célula de transcribir el gen que codifica el ADN afectado. Otras lesiones producen mutaciones potencialmente nocivas en el genoma de la célula, lo que afecta la supervivencia de sus «células hijas» a la hora de la mitosis. Por consiguiente, el proceso de reparación del ADN es constantemente activo, respondiendo a daños a la estructura del ADN.
 La velocidad de la reparación del ADN depende de muchos factores, como el tipo de célula, su edad, y el ambiente extracelular. Una célula que haya acumulado una gran cantidad de daños en el ADN, o que no pueda reparar eficazmente los daños producidos en su ADN, puede entrar en uno de tres estados posibles:
La velocidad de la reparación del ADN depende de muchos factores, como el tipo de célula, su edad, y el ambiente extracelular. Una célula que haya acumulado una gran cantidad de daños en el ADN, o que no pueda reparar eficazmente los daños producidos en su ADN, puede entrar en uno de tres estados posibles:- Un estado irreversible de inactividad, llamado senescencia.
- Suicidio celular, llamado apoptosis o muerte celular programada.
- Carcinogénesis, o formación de cáncer.
La capacidad de reparación del ADN es vital para la integridad de su genoma, y por tanto, de su funcionamiento normal y el del organismo. En el caso de muchos de los genes que se había demostrado que influían en la longevidad, más tarde se ha revelado que tienen un papel en la reparación y protección del ADN. La incapacidad de corregir lesiones moleculares en las células que forman gametos pueden introducir mutaciones en el genoma de sus descendientes, influyendo en el ritmo de la evolución.
Reparación por escisión de nucleótido (NER), que repara daños que afecten cadenas más largas, de entre dos y treinta bases. Este proceso reconoce cambios grandes que distorsionan la hélice, como dímeros de timina, así como roturas de cadena única (reparados con enzimas como la UvrABC endonucleasa. Una forma especializada de NER, conocida como reparación acoplada a transcripción (TCR) desarrolla enzimas de alta prioridad en genes que se están transcribiendo activamente. se corta la cadena de azúcar - fosfato. Los genes implicados, en el caso de
E. coli, son Uvr (A, B, C, D). Los mutantes en genes Uvr son muy
sensibles a la luz ultravioleta, por tanto, los genes Uvr confieren resistencia
a UV.

Reparación por escisión de base (BER), que repara daños a un único nucleótido causados por oxidación, alquilación, hidrólisis o desaminación. Una glicosidasa escinde la base nitrogenada del nucleótido dañado, generando un sitio apurínico o apirimidínico. El esqueleto pentosa-fosfato residual es eliminado por una AP endonucleasa y finalmente es sustituido por el nucleótido adecuado por la actividad secuencial de ADN polimerasa y ADN ligasa.

Reparación directa, no requiere eliminación de nucleótidos o bases nitrogenadas, sino que se emplean enzimas para reparar directamente alteraciones nucleotídicas. Los principales enzimas empleados son la fotoliasa (separa los dímeros de timinas formados por radiación UV) y la metiltransferasa (retira grupos metilo añadidos al ADN).
Reparación de malapareamiento o reparación por mismatch (MMR). Todas las reparaciones anteriores se realizan antes de terminar la replicación. Este sistema se realiza cuando la replicación ya ha concluido, y corrige errores de nucleótidos mal apareados (pero normales, es decir, no dañados). Para ello debe reconocer qué hebra es la correcta, lo que en procariotas ocurre porque el ADN suele tener metiladas sus bases, pero tras la replicación la hebra nueva no se metila hasta comprobar que no tenga errores, por lo que la maquinaria de reparación supone que si hay un error tras la replicación, se habrá producido en la hebra nueva (la no metilada). Una vez metiladas, o no hay corrección posible, o ésta puede causar errores. Por ejemplo, en cualquier emparejamiento erróneo de GT y CT, se retira preferentemente la timina, porque es probable que sea resultado de la desaminación de la citosina. Este sistema de reconocimiento por metilación solo funciona en procariotas, se ignora cuál es el mecanismo empleado en eucariotas para distinguir la hebra recién formada de la hebra madre.
 Respuesta SOS, que es un rellenado de emergencia que se pone en marcha cuando se acumulan daños que distorsionan la doble hélice (como regiones de ADN monocatenario, por pérdidas de nucleótidos en la cadena complementaria), atascando la maquinaria replicativa. En procariotas esto desencadena el sistema RecA, una proteasa que elimina proteínas represoras de polimerasas de bypass, capaces de sobreponerse a la distorsión de la hélice y rellenar los huecos con nucleótidos al azar. En eucariotas las polimerasas de bypass son constitutivas, están presentes en todo momento en el citoplasma, pero solo se reclutan cuando se acumulan daños, mediante una regulación por ubiquitinación de la abrazadera.
Respuesta SOS, que es un rellenado de emergencia que se pone en marcha cuando se acumulan daños que distorsionan la doble hélice (como regiones de ADN monocatenario, por pérdidas de nucleótidos en la cadena complementaria), atascando la maquinaria replicativa. En procariotas esto desencadena el sistema RecA, una proteasa que elimina proteínas represoras de polimerasas de bypass, capaces de sobreponerse a la distorsión de la hélice y rellenar los huecos con nucleótidos al azar. En eucariotas las polimerasas de bypass son constitutivas, están presentes en todo momento en el citoplasma, pero solo se reclutan cuando se acumulan daños, mediante una regulación por ubiquitinación de la abrazadera.
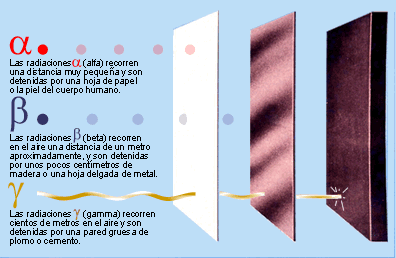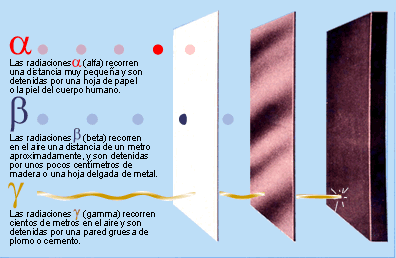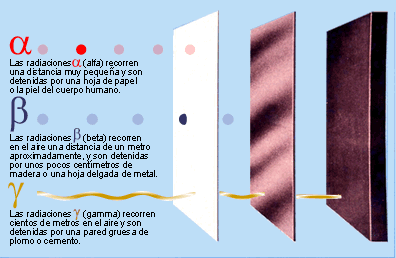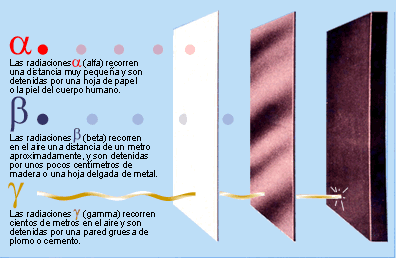
Los agentes que causan daños en el
DNA tienen distintos orígenes. Por una parte, están las alquilaciones
(metilaciones principalmente), las desaminaciones, la oxidación y los rayos UV.
La acumulación de mutaciones en células somáticas es el origen de muchos
cánceres y las células cancerosas reparan mal las mutaciones. Por eso muchos
tratamientos antineoplásicos sobre la base de inducir mutaciones que los
tumores no saben reparar.